天津科研技術服務分離
痙攣型腦性癱瘓(SCP)大鼠模型【簡介】痙攣性腦癱指發育異常引起的運動和感覺功能落后,造成肢體肌張力增高、腱反射亢進等一系列綜合征。本實驗利用腦立體定位解剖圖譜,對大鼠的錐體束部位進行精確的立體定位,通過微量注射器對大腦錐體束所在部位注射無水乙醇造成錐體束壞死,的模擬了痙攣型腦癱的解剖學及病理改變,術后大鼠產生明顯的屈曲痙攣癥狀,且屈肌肌張力增高,癥狀和體持續時間較長,穩定性良好。從而建立一種可復制性良好且痙攣持續時間較長的穩定的大鼠痙攣型腦癱動物模型,為進一步深入的探究痙攣型腦癱的基礎研究和臨床診治打下基礎【目的】痙攣型腦性癱瘓(SCP)大鼠模型建立【動物】SPF級SD大鼠,雄性,周齡6~8W,體重:200~250g【方法】1、大鼠麻醉,顱頂脫毛備皮;2、大鼠腦定位儀固定大鼠,顱頂正中切口,長度約2cm開口暴露前囟及矢狀縫;3、縫線將皮膚左右分開固定,前囟后10mm、矢狀縫左側;4、微量注射器移至開孔處修正骨孔,注射器垂直向顱內插入,緩慢注射無水乙醇15ul,注射完移除注射器,棉球壓迫止血;5、縫合創口后維持25°體溫,等待蘇醒,觀察大鼠狀態。【觀察】術后3天模型大鼠攝食減少、右側肢體跛行、右前肢不負重、右前肢及右前爪屈曲痙攣明顯。干貨分享 | 瓊脂糖核酸電泳,選擇紫外還是藍光?天津科研技術服務分離

圖2MAZTER-Seq實驗流程圖圖3MAZTER-MINE分析m6A示意圖接下來作者便是要驗證這一新方法的可行性了。在酵母中敲除IME4的情況下,檢測到的剪切效率高于野生型(剪切效率高低m6A水平),m6A抗體富集后的樣品剪切效率也低于未富集的Input組。整體水平可靠,那檢測的特異性位點是否準確呢?作者也將該方法檢測到的新甲基化位點使用放射標記層析檢測,發現預測的位點準確存在而且與剪切效率相符合。如圖5所示。而圖6中,作者則是與m6A抗體IP的方法進行了比較,也證實了這一方法的可行性。圖5MAZTER-Seq檢測結果驗證圖6MAZTER-Seq與m6A-Seq比較分析此外,后文中作者也在大規模的CRISPR-Cas9改變m6A狀態和酵母減數分裂模型中檢測了MAZTER-Seq這一系統;并進一步通過這一方法檢測了哺乳動物不同細胞間m6A水平的保守性;也探究了去甲基化酶FTO對整體m6A甲基化水平的影響等。這里小編主要給大家分享這一新技術,其他部分暫不過多分析了。新的技術能拓展我們的研究內容;對于這一技術。天津疾病科研技術服務外包動物疾病模型可以分為兩種類型:自然模型和人工模型。
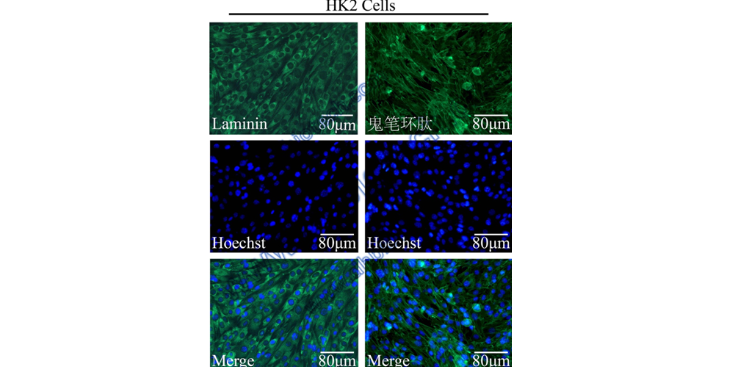
3)基因/蛋白質表達定性或定量檢測在進行分子檢測的過程中,保持核酸和蛋白質的完整性至關重要,因此在取樣過程中,應盡量避免核酸及蛋白質的降解。如不注意采樣過程,將會導致樣本中的核酸及蛋白質的無差別降解,嚴重影響后續分子檢測結果。在條件允許的情況下,樣本在離體后應立刻裝入凍存管內,并立即放入液氮中進行冷凍。在RNA提取樣本的保存過程中,可以選擇非冷凍型RNA保存液或Trizol試劑進行RNA酶的。針對蛋白質提取樣本的保存,我們同樣可以使用商品化的蛋白酶劑進行短期處理。聚合酶鏈式反應(PolymeraseChnReaction,PCR)及免印跡(Westernblotting,WB),這兩種實驗手段是分子學檢測中不可或缺的一部分,也是發表至關重要的分子檢測數據。PCR主要用來對基因在基因組層面或轉錄層面的變化進行定性或定量檢測,而WB則主要用來對某一蛋白質的表達進行定量檢測。(4)轉錄組學、蛋白質組學及代謝組學檢測隨著檢測水平及相關產業的不斷發展,各類組學檢測的價格降低,實驗設計中也常常運用組學檢測來提升實驗設計的檔次。在我們進行轉錄組學及蛋白質組學的檢測過程中,同樣是要首先確保目標RNA或蛋白質的片段完整性。
用途基因功能研究、免/殺傷/增殖等、抗體活性篩選(細胞水平的結合和阻斷)、CAR分子的殺傷活性評價。材料與儀器(以慢pMSCV載體為例)包含目的基因和eGFP-Tag的pMSCV質粒、帶有eGFP-Tag的pMSCV空載質粒、GAG質粒、VSV質粒、Puromycin、Polybrene、Lipo3000、HEK293T細胞、opti-MEM、DMEM、FBS、雙抗、μm的濾膜、熒光顯微鏡等。步驟1、基因的構建1)根據目的基因mRNA編碼區設計引物,分別在引物兩端加入酶切位點EcoRI和BglII。2)從細胞中提取目的基因mRNA,然后逆轉錄成cDNA,然后從cDNA里面用引物把目的基因的CDS區擴增出來。PCR擴增出帶有酶切位點的目的基因編碼區序列,連接至pMD19-T載體后轉化至感受態DH5α,分別進行菌落PCR鑒定和酶切鑒定。3)使用EcoRI和BglII雙酶切下目的基因序列,電泳割膠回收純化,連接到pMSCV-eGFP載體。再次轉化到感受態DH5α,菌落PCR鑒定和酶切鑒定成功后,送至公司測序、鑒定。4)鑒定成功后,將質粒轉化至感受態DH5α中,并進行無內質粒抽提。GAG質粒和VSV質粒同樣可以轉化至感受態DH5α中,并進行無內質粒抽提。質粒抽提后冷凍于-20℃保存。2、慢包裝1)使用DMEM完全培養基培養6cm皿HEK293T至匯合度為70~80%。由于大部分研究使用到的是人類來源的細胞,種植到免疫正常的動物體內會發生免疫排斥反應。
外泌體的抑制作用外泌體水平升高通常與不同類型的惡化相關,一些研究人員希望能通過降低外泌體到正常水平來防止不良預后。從這個角度出發,許多正在進行的研究旨在通過調節外泌體生成分泌的過程或通過特異性靶向其成分抑制其與靶細胞的相互作用來調節外泌體的產生。如今我們對外泌體機制和不同生理和病理條件下的功能的理解呈指數級增長,雖然目前其生物學功能還未完全解析清楚,但研究者們在許多領域均已對其進行了深入探索。外泌體不是廢物顆粒,而是細胞間通訊的關鍵介質,作為宿主細胞的衛星,外泌體包含大量的生物信息,其功能超出了初的預期,宿主細胞控制著外泌體的內容物,從而改變了自己或其他細胞的命運。其次,外泌體對的進展和轉移有著強烈的影響,可以通過外泌體預測轉移的部位并建立轉移前的生態位。參考文獻:[1]高方園,焦豐龍,張養軍,秦偉捷,錢小紅.外泌體分離技術及其臨床應用研究進展[J].色譜,2019,37。科研小白如何摸索自己的預實驗。江蘇乳鼠科研技術服務構建
動物模型的表現與人類疾病可能存在差異,因此需要謹慎使用。天津科研技術服務分離
且研究表明HNRNPC通過m6A與RNA結合調控目標轉錄本的豐度和選擇性剪切[9].圖1m6A修飾的酶系統[10]m6A生物學功能越來越多的證據表明m6A修飾在哺乳動物中發揮重要的生物功能。例如,在轉錄水平上調控RNA的穩定性[11]、定位[12]、運輸、剪切[13]和翻譯[14]。ClaudioR.等發現依賴METTL3的pri-miRNA甲基化,會促進DGCR8識別和加工,從而促進microRNA的成熟[15]。此外,m6A識別蛋白HNRNPA2B1促進pri-miRNA加工成pre-miRNA[16]。另外,環狀RNA上m6A的修飾能促進環狀RNA的翻譯[17]。m6A修飾在基因表達調控中起著重要的作用,其調控機制的異常可能與人類疾病或相關。目前發現m6A可能會影響精子發育(ALKBH5,METTL3,Ythdc2)、發育(METTL3、FTO、ALKBH5)、免疫(METTL3)、UV誘導的DNA損傷反應(METTL3,FTO)、生成(YTHDF2)或轉移(METTL14)、干細胞更新(METTL14)、脂肪分化(FTO)、生物節律、細胞發育分化、細胞分裂及其它的一些生命過程。例如,ALKBH5敲除的雄性小鼠增加了mRNA中的m(6)A修飾,其特點是凋亡影響減數分裂中期的精子細胞,引起生育能力受損[7]。METTL3和METTL14增加弱精癥精子的m6A水平[18],在生殖細胞中,METTL3的敲除嚴重抑制精子分化和減數分裂的發生。天津科研技術服務分離
- 寧夏裸鼠原代細胞技術 2025-12-06
- 吉林血液原代細胞 2025-12-06
- 上海組織科研技術服務培養 2025-12-06
- 湖北血液科研技術服務技術 2025-12-06
- 基因敲除動物模型造模 2025-12-06
- 湖北哪里有科研技術服務技術 2025-12-06
- 模式原代細胞外包 2025-12-06
- 山西實驗科研技術服務技術 2025-12-06
- 江蘇模式原代細胞構建 2025-12-06
- 模式原代細胞服務 2025-12-06
- 青浦區一站式文化藝術活動策劃平臺 2025-12-06
- 杭州拱墅區尋釁滋事刑事辯護監外執行 2025-12-06
- 奉賢區提供舞臺設備銷售便捷 2025-12-06
- 泉州機加工勞務派遣公司推薦 2025-12-06
- 太原創新型企業服務中心數實融合趨勢 2025-12-06
- 吉林臻推寶多少錢 2025-12-06
- 徐州哪些PTI試驗粉塵 2025-12-06
- 新吳區本地懸掛式七氟丙烷滅火消防裝置 2025-12-06
- 工業區合同節水是什么 2025-12-06
- 過程追溯業務 2025-12-06